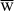 s = average fitness of non altruistsa = average fitness of altruists
s = average fitness of non altruistsa = average fitness of altruistsGeneral rule for the evolution of altruism.
Let:
s = average fitness of non altruistsa = average fitness of altruistsWe calculate the mean fitness of selfish individuals and altruists as:
s = wo + Bps.
a = wo + Bpa - C
Bps = Contribution of altruists to the fitness of selfish individuals.
Bpa = Contribution of altruists to the fitness of other altruists.
Altruism can increase when a > s, meaning:
B(pa-ps) > C
The term (pa-ps) is the expected amount of altruism experienced by an altruist, and the expected amount experienced by a non-altruist.
This will, on average, be zero if altruists interact randomly with others in a very large population.
To understand the evolution of altruism, we need to understand the ways in which (pa-ps) can become positive.
We have already seen one way to increase (pa-ps).
If individuals are most altruistic towards others that have helped them in the past, pa goes up and ps decreases.
Recall that this was called reciprocal altruism.
Another way to increase (pa-ps) is for altruists to direct their attention to close relatives, since these are likely to also have alleles for altruism.
This is called Kin Selection
It is common to see (pa-ps) written as a single term: 'r'.
The condition for altruism to increase in frequency then becomes:
Br > C
This is Hamilton's Rule for the evolution of altruism.
The symbol 'r' was originally used because the value (pa-ps) is sometimes well approximated by the genealogical relatedness between individuals.
The meaning of r:
For a rare allele in a very large population, the probability that an individual who does not have the allele encounters one who does is approximately zero. If you have such an allele, the probability that another individual also has it is almost exactly the probability that they and you both inherited it from a recent common ancestor.
(ignoring the prob. that it just arose by mutation and assuming that because it is rare, it is quite unlikely that both parents had it or that anyone was homozygous)
For full siblings:
r = P[rare allele came from mother]*P[sibling got same allele from mother]
+ P[rare allele came from father]*P[sibling got same allele from father]
For diploid organisms, each offspring gets 1/2 of each parent's genes. We can find the value of r using a path diagram:
With parent r = 1/2
Full siblings r = 1/2 (= 1/2 * 1/2 + 1/2 * 1/2)
Half sibs r = 1/4 (= 1/2 * 1/2)
First cousins r = 1/8
Thus, a rare allele that caused individuals carrying it to sacrifice their lives to save the lives of more than two full siblings would increase in frequency in the population (if the population were structured as above).
Example: Cannibalism in Amphibians.
Among some toads and salamanders, juveniles may
develop into either omnivores or obligate carnivores, the latter often
eating members of the same species.
In both spadefoot toads and tiger salamanders, cannibalistic individuals are significantly less likely to eat close relatives
than those to whom they are not related.
Cannibalistic tiger salamanders distinguish between degrees of
relatedness, eating cousins more often than siblings, and unrelated
individuals more often than cousins.
Eusociality:
Situation in which some individuals give up reproduction altogether
in order to help others raise offspring.
Has arisen in:
Hymenopteran insects (ants, bees, wasps) - 9 times
Aphids - many times (> 5)
Termites - 1 time
Ambrosia Beetles - 1 time
Thrips - 1 time
Snapping Shrimp - 3 times
Mammals - 2 time (Naked Mole Rats and Damaraland Mole Rats)
Hymenopterans are notable in that the sterile "workers" are all females (in other eusocial groups the sterile castes include both males and females).
Hymenopteran insects
Have a haplodiploid sex determining system:
Haploid = male
Diploid = female
Among full sib sisters, half of their genome is from their father, who
is haploid and thus passed all of his genes to each daughter.
The path diagram from a female to her full sib sister thus looks like this:
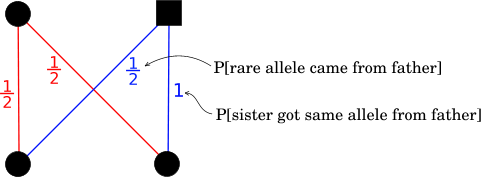
Thus, for full sib sisters:
P[share rare allele from mother] = 1/2 * 1/2 = 1/4
P[share rare allele from father] = 1/2 * 1 = 1/2
So: r = 3/4
For sisters relative to brothers:
P[allele in sister also found in brother] = 1/2 * 1/2 = 1/4
P[allele in brother also found in sister] = 1 * 1/2 = 1/2
For mother/offspring, r = 1/2
For father/offspring, r = 1
This makes sense of the fact that male hymenopterans are not workers.
Males more closely related to their own offspring (r = 1) than to their siblings (r = 1/2).
Note that females are related to their siblings, on average, by 1/2 (3/4 for sisters, 1/4 for brothers).
Multilevel Selection
Just as the efficacy of selection on individuals is determined by the
amount of phenotypic variation between them, the strength of selection
between groups is determined by the amount of variation between groups.
If within group variation is high but between group variation is low, then selection at the level of individual organisms tends to drive evolution.
If within group variation is low but between group variation is large,
then selection at the level of groups is the more effective evolutionary
force.
This tends to maximize overall group reproductive output.
Example: Sex ratio in Fig Wasps
In some organisms, one or a few females lay eggs in a resource patch
(such as a fig blossom for fig wasps), but when the eggs hatch, the offspring
stay in the patch.
In fig wasps, the offspring grow up inside the developing fig and mate there.
After mating, females leave the fig and disperse to lay eggs in another fig.
In such cases, we generally see a female biased sex ratio.
Explanation : Within a fig, selection favors a 50/50 sex ratio, in accordance with Fisher's theory.
However, those groups that have a female biased sex ratio produce more descendants to disperse to form new colonies.

Thus, selection at the colony level favors those colonies that have a female biased sex ratio.
This only works because the colonies are temporarily isolated from one
another.
If they were not, then males would simply move into those groups that have
a preponderance of females.
We would expect this effect to be strongest when there is maximal variation between groups (figs) and minimal variation within each fig.
This is what happens when only one female lays eggs in a given fig.
As the number of females laying eggs in each fig increases, the variation within figs increases and the variation between them decreases.
Different types of fig wasp differ in how many females lay in a fig.
We thus expect that the sex ratio will be closer to 1:1 for species in which many females lay eggs in each fig , and more female biased as the number of foundresses per fig declines.
This is exactly the pattern that is seen.
