
a) On graph (A): Draw a fitness function for another strategy, S2, such that there are two pure strategy ESS's.
b) On graph (B): Draw a fitness function for S2 that creates a single mixed ESS at Freq(S1) = 0.3. (5 pts.)
Review vocabulary for Exam 3
Species selection
Bdelloid rotifers
Heritability
Additive genetic variance
Selection differential
Response to selection
Breeder's equation
Price equation
Additive genetic covariance
Fitness gradient vector
G matrix
Allometry
Heterochrony
Progenesis
Hypermorphosis
Neoteny
Acceleration
Sexual selection
Isogamy
Anisogamy
Runaway sexual selection
Indicator trait
Frequency dependant selection
Evolutionarily stable strategy
Altruism
Prisoners' dilemma
Tit-for-tat
Win-stay-lose-shift
Reciprocal altruism
Eusociality
Hamilton's rule
Kin selection
Multilevel selection
Some review questions for Exam 3
1) Imagine a storm that kills many insects, sparing principally those with short wings such that the average wing length of survivors is 2 mm shorter than the average before the storm. The survivors reproduce and their offspring have wings that are, on average, .5 mm longer than those of their parents. What is the value of h2 for this character?
2 The example in the previous question is based on a real case, in which it was also observed that the average thorax length among the survivors was slightly less than that in the population before the storm. Other evidence, however, indicates that a long thorax helps these insects survive strong winds. Explain how thorax length could decreased in spite of selection for it to increase.
3) What would be the value of 'r' between workers in a hymenopteran colony if the queen had mated with two different males?
4) It is often difficult to measure selection directly unless you already have individuals marked and measured. However, there are some values that we can often measure: 1) The change in the mean value of some trait over a generation, and 2) the heritability of the trait. Explain how you could use these pieces of information to calculate the selection differential.
5) How could you design a selection experiment to estimate additive genetic covariance?
6) What are the two main mechanisms for the evolution of extreme male traits? Explain what role(s) genetic covariance plays in these mechanisms.
7)In what sense were the antlers of Irish Elk just the size that we should expect them to be, while the antlers of Moose are smaller than expected?
8)In what way was the evolution of anisogamy a necessary prerequisite to the evolution of sexual dimorphism?
9)Why is it that the skeletons of large vertebrates, on the whole, do not support their bodies (on land) as well as do those of small vertebrates?
10) In some organisms, individuals do not mate randomly but rather form local groups that persist for a couple of generations, after which the resulting progeny disperse to form new groups. In such cases, we often see a deviation from the common 1:1 sex ratio. In what way do you expect the sex ratio in such cases to differ from 1:1? Why is it different from what we see in randomly mating populations?
11) Consider the fitness functions
for strategy S1 shown here
(both are the same).
a) On graph (A): Draw a fitness function for another strategy, S2, such that there are two pure strategy ESS's.
b) On graph (B): Draw a fitness function for S2 that creates a single mixed ESS at Freq(S1) =
0.3. (5 pts.)
12) Describe how kin selection and reciprocal altruism can be described as variants of the same basic principle underlying the evolution of altruism.
13) Explain why, under directional selection, the variance in a quantitative trait does not go down as the mean changes. Where does the new variation come from?
Answers and Hints
1)Here, S = -2 and R = -1.5. Rearranging the equation R = h2 S yields h2 = R/S. So h2 = 3/4.
2)Thorax length and wing length are probably genetically correlated. In such a case, selection on one will lead to change in the other as well.
3)
Prob(allele came from mother) = .5
Prob(sister got same allele if it came from mother) = .5
Prob(allele came from father) = .5
Prob(sister has the same father) = .5
r = .5*.5 + .5*.5 = .5
4) The change in mean value of the trait = R
Heritability is h2
Now use R = h2S to find S.
5) Select on one character (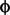 1) and observe the change in the other
(making sure that it is not under selection). the change in the non selected
character (2) should be:
1) and observe the change in the other
(making sure that it is not under selection). the change in the non selected
character (2) should be:
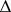 2 = 1/
2 = 1/ CovA(1,
2)
CovA(1,
2) ![]() w,1
w,1
Since you imposed the selection, you can calculate CovA(1,
2).
6) Runaway sexual selection, and selection for indicator traits.
b) Runaway sexual selection involves the joint increase in frequency of a trait (that reduces survivorship) in one sex and the preference for that trait in the other sex. This can occur only if there is an additive genetic covariance between the trait and the preference.
Indicator models describe cases in which females prefer to mate with males that express some trait (which may or may not reduce survivorship) because that trait covaries with some other, unobservable, trait that is advantageous. In this case, the covariance that matters is between two traits expressed in the same sex.
7)The antlers of the Irish Elk lie almost exactly along the allometric line for all deer. Thus, though they are larger in absolute size than those of other species, they are just the size that we would expect them to be for a deer of that size. By contrast, the antlers of Moose lie well below the allometric line, meaning that they are smaller than we would expect given the Moose's body size.
8)Anisogamy, the condition in which some gametes are large (e.g. eggs) and others small (e.g. sperm) created a situation in which the optimal reproductive strategies are different for individuals that produce the large gametes than for those who produce the small gametes. In particular, since fewer large gametes can be produced by a given individual, those individuals tend to invest more in each offspring an put a lower emphasis on producing very many offspring than do the individuals that produce small gametes.
9)The strength of a bone is proportional to it's cross sectional area. Since the overall mass of the animal is proportional to length3, the cross sectional area of the bones must also scale as length3 if they are to be equally good at supporting the animal. The mass of the bone is proportional to it's length times it's cross sectional area, so the overall mass of bones must scale as length4, or mass4/3, in order to be equally supportive. At this rate, a large terrestrial vertebrate would have to be nearly all skeleton. In fact, bone mass scales as body mass to the 1.083 power, which means that large vertebrates have proportionally less supportive skeletons.
10)These conditions are right for selection to act between these groups. Selection between groups favors those that have the highest reproductive output (mean fitness), so this will select for a female biased sex ratio. If the population is not structured in this way (individuals mate randomly), then we expect a 1:1 sex ratio as this is an ESS.
11)

12) Altruism can increase in frequency if B(pa - ps) > C, where B is the benefit to the recipient of an altruistic act, C is the cost to the altruist, and (pa - ps) is the degree to which altruism is preferentially directed at altruists. Both kin selection and reciprocal altruism involve increasing the value of (pa - ps). In kin selection, this happens because altruists preferentially help relatives, who are more likely to have alleles for altruism than a random individual would be. In reciprocal altruism, (pa - ps) is increased because altruists preferentially help others who, by their past actions, have shown themselves to also be altruists.
13) Sexual reproduction reshuffles alleles every generation. When directional selection is increasing the frequencies of formerly rare alleles, this reshuffling produces many new genotypes, adding new variation to the population every generation.
======================================================
A note on locating ESS points: An equilibrium is just a point with the property that if the system is exactly at that point, it stays there. When we are studying evolution by following the frequency of something, the points 0 and 1 are always equilibria.
As a rule, the equilibria are at 0, 1, and any point at which the fitness lines cross (if such a point exists).
We determine whether or not an equilibrium is stable by looking at points that are close to it. If, from nearby points, the system moves towards the equilibrium, it is stable. If, from nearby points, the system moves away, then the equilibrium is unstable.